La Fotobiología es la ciencia que estudia el efecto de la radiación no ionizante sobre los sistemas vivos. Se hace una revisión de los conceptos básicos relativos a la luz, a la materia y la interacción de la radiación lumínica y materia que están en el campo de la biofísica.
FENÓMENOS FOTOQUÍMICOS.
La fotoquímica es el estudio de las transformaciones químicas provocadas o catalizadas por la emisión o absorción de luz visible o radiación ultravioleta. Una molécula en su estado fundamental (no excitada) puede absorber un quantum de energía lumínica, esto produce una transición electrónica y la molécula pasa a un estado de mayor energía o estado excitado. Una molécula excitada es más reactiva que una molécula en su estado fundamental.
El fenómeno fotoquímico precisa de fases principales:
1.Recepción de la energía luminosa.
FENÓMENOS FOTOQUÍMICOS.
La fotoquímica es el estudio de las transformaciones químicas provocadas o catalizadas por la emisión o absorción de luz visible o radiación ultravioleta. Una molécula en su estado fundamental (no excitada) puede absorber un quantum de energía lumínica, esto produce una transición electrónica y la molécula pasa a un estado de mayor energía o estado excitado. Una molécula excitada es más reactiva que una molécula en su estado fundamental.
El fenómeno fotoquímico precisa de fases principales:
1.Recepción de la energía luminosa.
2.Reacción química propiamente dicha.
Según se opere con una sustancia única o con un sistema de varios cuerpos en presencia, se realizará, bien una descomposición de la sustancia en sus elementos (fotólisis), bien una combinación de varios cuerpos en uno solo (fotosíntesis).
Leyes fundamentales.
1. Ley de absorción de Grotthus-Draper: Una radiación no puede provocar acción química más que si es absorbida por un cuerpo (o un sistema de cuerpos); si no, no puede haber transmisión de energía luminosa.
Es conveniente señalar que las radiaciones que constituyen el color de un cuerpo son justamente las no absorbidas. No tienen, por lo tanto, efecto sobre el mismo. Por el contrario las radiaciones complementarias de éste color son absorbidas y son susceptibles de acción. Por ejemplo, una sustancia de color verde emite el verde pero absorbe el rojo y el azul. No podrá ser descompuesta más que por estos dos últimos colores.
2. Ley energética: Para que una radiación luminosa actúe eficazmente, debe poseer una energía, por lo menos, igual a la necesaria para la transformación química.
Se sabe que la radiaciones poseen tanta más energía cuanto más cortas sean sus longitudes de onda (o más elevadas sean sus frecuencias). La energía transportada por un fotón viene dada por la expresión:
E= hv = (hc)/ phi
( c= velocidad de la luz)
en la que h es la constante de Plank, igual a 6,55x10-27 ergios.
3. Ley de la equivalencia fotoquímica (o ley de Einstein): A cada fotón absorbido, corresponde una molécula descompuesta o combinada.
P= MOLÉCULAS DESCOMPUESTAS/ Nº DE FOTONES ABSORBIDOS.
Se sobreentiende que los fotones activos satisfacen la ley energética precedente.
Según esto se comprueba que prácticamente el número de fotones activos absorbidos en una reacción química, corresponde raramente al número de moléculas descompuestas con el número de fotones absorbidos, se obtiene un rendimiento cuántico que varía entre amplios límites, de 0,1 a 1000 (y más). Sólo algunas reacciones tienen un rendimiento teórico igual a la unidad.
A pesar de estas contradicciones, no se puede poner en duda la validez de la ley de Einstein, y la razón de las variaciones experimentales es simple:
a) Cuando la reacción química exige una aportación de energía (reacción endotérmica), como en el caso de los haluros de plata, r es todo lo más igual a 1. En general es más pequeño, como en la descomposición fotoquímica del clorhídrico gas, pues esta reacción es reversible.
Para descomponer el amoníaco NH3 en nitrógeno e hidrógeno, por los rayos ultravioletas, son precisos cuatro fotones por molécula (= 0,25.).
Según la longitud de onda, se puede modificar el equilibrio fotoquímica a un sentido u otro. Así, en la reacción reversible ácido maléico ácido fumárico donde con el ultravioleta = 313 mm existe 44 % de ácido maleico y 56 % de ácido fumárico, mientras que con una onda más corta = 200 mm, el ácido maleico se regenera, con un 75 % de ácido maleico y 25 % de ácido fumárico. En el primer caso, el rendimiento cuántico es de 0,03 mientras que se eleva a 0,1, por la reacción inversa.
b) Cuando las radiaciones absorbidas provocan primero una activación de la molécula, que reacciona a continuación sobre una segunda molécula neutra para dar productos de descomposición, según el esquema siguiente:
AB + hv = (AB)
(AB) + AB = 2A + 2B
el rendimiento cuántico es casi igual a 2.
Se sobreentiende que los fotones activos satisfacen la ley energética precedente.
Según esto se comprueba que prácticamente el número de fotones activos absorbidos en una reacción química, corresponde raramente al número de moléculas descompuestas con el número de fotones absorbidos, se obtiene un rendimiento cuántico que varía entre amplios límites, de 0,1 a 1000 (y más). Sólo algunas reacciones tienen un rendimiento teórico igual a la unidad.
A pesar de estas contradicciones, no se puede poner en duda la validez de la ley de Einstein, y la razón de las variaciones experimentales es simple:
a) Cuando la reacción química exige una aportación de energía (reacción endotérmica), como en el caso de los haluros de plata, r es todo lo más igual a 1. En general es más pequeño, como en la descomposición fotoquímica del clorhídrico gas, pues esta reacción es reversible.
Para descomponer el amoníaco NH3 en nitrógeno e hidrógeno, por los rayos ultravioletas, son precisos cuatro fotones por molécula (= 0,25.).
Según la longitud de onda, se puede modificar el equilibrio fotoquímica a un sentido u otro. Así, en la reacción reversible ácido maléico ácido fumárico donde con el ultravioleta = 313 mm existe 44 % de ácido maleico y 56 % de ácido fumárico, mientras que con una onda más corta = 200 mm, el ácido maleico se regenera, con un 75 % de ácido maleico y 25 % de ácido fumárico. En el primer caso, el rendimiento cuántico es de 0,03 mientras que se eleva a 0,1, por la reacción inversa.
b) Cuando las radiaciones absorbidas provocan primero una activación de la molécula, que reacciona a continuación sobre una segunda molécula neutra para dar productos de descomposición, según el esquema siguiente:
AB + hv = (AB)
(AB) + AB = 2A + 2B
el rendimiento cuántico es casi igual a 2.
FOTOSÍNTESIS DESDE EL PUNTO DE VISTA DE LOS PIGMENTOS Y CAPTACIÓN DE LA LUZ.
La fotosíntesis es la conversión de energía luminosa en energía química estable, siendo el adenosín trifosfato (ATP) la primera molécula en la que queda almacenada esa energía química.
Con posterioridad, el ATP se usa para sintetizar moléculas orgánicas de mayor estabilidad.
Los orgánulos citoplasmáticos encargados de la realización de la fotosíntesis son los cloroplastos, unas estructuras polimorfas y de color verde (esta coloración es debida a la presencia del pigmento clorofila) propias de las células vegetales. En el interior de estos orgánulos se halla una cámara que contiene un medio interno llamado estroma, que alberga diversos componentes, entre los que cabe destacar enzimas encargadas de la transformación del dióxido de carbono en materia orgánica y unos sáculos aplastados denominados tilacoides o lamelas, cuya membrana contiene pigmentos fotosintéticos. En términos medios, una célula foliar tiene entre cincuenta y sesenta cloroplastos en su interior.
La energía luminosa que absorbe la clorofila se transmite a los electrones externos de la molécula, los cuales escapan de la misma y producen una especie de corriente eléctrica en el interior del cloroplasto al incorporarse a la cadena de transporte de electrones. Esta energía puede ser empleada en la síntesis de ATP mediante la fotofosforilación, y en la síntesis de NADPH. Ambos compuestos son necesarios para la siguiente fase o Ciclo de Calvin, donde se sintetizarán los primeros azúcares que servirán para la producción de sacarosa y almidón. Los electrones que ceden las clorofilas son repuestos mediante la oxidación del H2O, proceso en el cual se genera el O2 que las plantas liberan a la atmósfera.
Existen dos variantes de fotofosforilación: acíclica y cíclica, según el tránsito que sigan los electrones a través de los fotosistemas. Las consecuencias de seguir un tipo u otro estriban principalmente en la producción o no de NADPH y en la liberación o no de O2.
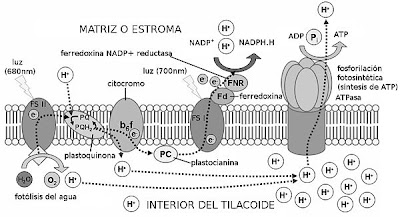 Fotofosforilación acíclica.
Fotofosforilación acíclica.
Este proceso permite la formación de ATP y la reducción de NADP+ a NADPH + H+, y necesita de la energía de la luz, como ya se ha dicho. Se realiza gracias a los llamados fotosistemas, que se encuentran en la membrana de los tilacoides (en los cloroplastos). Estos están formados por dos partes:
Antena, donde se agrupan los pigmentos antena, junto con proteínas, y cuya función es captar la energía de los fotones para transmitirla al pigmento diana; y el centro de reacción. Este está formado por proteínas y por pigmentos, encontrándose en él el llamado pigmento diana, que es aquel que recibe la energía de excitación de la antena, energía que sirve para excitar y liberar electrones. Aquí también se encuentra el primer dador de electrones, que repone los electrones al pigmento diana,
Primer aceptor, que recibe los electrones liberados.
Hay dos tipos de fotosistemas:
Fotosistema I, que se encuentra sobre todo en los tilacoides de estroma, y cuyo pigmento diana es la clorofila P700.
Fotosistema II, que se encuentra sobre todo en los grana y cuyo pigmento diana es la clorofila P680.
Proceso.
El proceso de la fase luminosa, supuesto para dos electrones, es el siguiente: Los fotones inciden sobre el fotosistema II, excitando y liberando dos electrones, que pasan al primer aceptor de electrones, la feofitina. Los electrones los repone el primer dador de electrones, el dador Z, con los electrones procedentes de la fotólisis del agua en el interior del tilacoide (la molécula de agua se divide en 2H+ + 2e- + 1/2O2). Los protones de la fotólisis se acumulan en el interior del tilacoide, y el oxígeno es liberado.
Los electrones pasan a una cadena de transporte de electrones, que invertirá su energía liberada en la síntesis de ATP. ¿Cómo? La teoría quimioosmótica nos lo explica de la siguiente manera: los electrones son cedidos a las plastoquinonas, las cuales captan también dos protones del estroma. Los electrones y los protones pasan al complejo de citocromos bf, que bombea los protones al interior del tilacoide. Se consigue así una gran concentración de protones en el tilacoide (entre éstos y los resultantes de la fotólisis del agua), que se compensa regresando al estroma a través de las proteínas ATP-sintasas, que invierten la energía del paso de los protones en sintetizar ATP. La síntesis de ATP en la fase fotoquímica se denomina fotofosforilación.
Los electrones de los citocromos pasan a la plastocianina, que los cede a su vez al fotosistema I. Con la energía de la luz, los electrones son de nuevo liberados y captados por el aceptor A0. De ahí pasan a través de una serie de filoquinonas hasta llegar a la ferredoxina. Ésta molécula los cede a la enzima NADP+-reductasa, que capta también dos protones del estroma. Con los dos protones y los dos electrones, reduce un NADP+ en NADPH + H+.
El balance final es: por cada molécula de agua (y por cada cuatro fotones) se forman media molécula de oxígeno, 1,3 moléculas de ATP, y un NADPH + H+.
Fase luminosa cíclica.
En la fase luminosa o fotoquímica cíclica interviene de forma exclusiva el fotosistema I, generándose un flujo o ciclo de electrones que en cada vuelta da lugar a síntesis de ATP. Al no intervenir el fotosistema II, no hay fotólisis del agua y, por ende, no se produce la reducción del NADP+ ni se desprende oxígeno. Únicamente se obtiene ATP.
El objetivo que tiene la fase cíclica tratada es el de subsanar el déficit de ATP obtenido en la fase acíclica para poder afrontar la fase oscura posterior.
Cuando se ilumina con luz de longitud de onda superior a 680 nm (lo que se llama rojo lejano) sólo se produce el proceso cíclico. Al incidir los fotones sobre el fotosistema I, la clorofila P700 libera los electrones que llegan a la ferredoxina, la cual los cede a un citocromo b6 y éste a la plastoquinona (PH), que capta dos protones y pasa a (PQH2). La plastoquinona reducida cede los dos electrones al citocromo f e introduce
Tiene lugar al mismo tiempo que la acíclica. En ella sólo interviene el fotosistema I. Los electrones liberados, después de llegar a la ferredoxina, pasan a las plastoquinonas, y siguen la cadena de transporte de electrones hasta regresar a la plastocianina y al fotosistema I. Por tanto, se genera ATP pero no NADPH. Sirve para compensar el hecho de que en la fotofosforilación acíclica no se genera suficiente ATP para la fase oscura.
BIBLIOGRAFÍA.
http://sisbib.unmsm.edu.pe/BVRevistas/dermatologia/v12_n2/fotobiologia_cutanea.htm
http://www.textoscientificos.com/fotografia/fotoquimica
Los orgánulos citoplasmáticos encargados de la realización de la fotosíntesis son los cloroplastos, unas estructuras polimorfas y de color verde (esta coloración es debida a la presencia del pigmento clorofila) propias de las células vegetales. En el interior de estos orgánulos se halla una cámara que contiene un medio interno llamado estroma, que alberga diversos componentes, entre los que cabe destacar enzimas encargadas de la transformación del dióxido de carbono en materia orgánica y unos sáculos aplastados denominados tilacoides o lamelas, cuya membrana contiene pigmentos fotosintéticos. En términos medios, una célula foliar tiene entre cincuenta y sesenta cloroplastos en su interior.
La energía luminosa que absorbe la clorofila se transmite a los electrones externos de la molécula, los cuales escapan de la misma y producen una especie de corriente eléctrica en el interior del cloroplasto al incorporarse a la cadena de transporte de electrones. Esta energía puede ser empleada en la síntesis de ATP mediante la fotofosforilación, y en la síntesis de NADPH. Ambos compuestos son necesarios para la siguiente fase o Ciclo de Calvin, donde se sintetizarán los primeros azúcares que servirán para la producción de sacarosa y almidón. Los electrones que ceden las clorofilas son repuestos mediante la oxidación del H2O, proceso en el cual se genera el O2 que las plantas liberan a la atmósfera.
Existen dos variantes de fotofosforilación: acíclica y cíclica, según el tránsito que sigan los electrones a través de los fotosistemas. Las consecuencias de seguir un tipo u otro estriban principalmente en la producción o no de NADPH y en la liberación o no de O2.
 Fotofosforilación acíclica.
Fotofosforilación acíclica.Este proceso permite la formación de ATP y la reducción de NADP+ a NADPH + H+, y necesita de la energía de la luz, como ya se ha dicho. Se realiza gracias a los llamados fotosistemas, que se encuentran en la membrana de los tilacoides (en los cloroplastos). Estos están formados por dos partes:
Antena, donde se agrupan los pigmentos antena, junto con proteínas, y cuya función es captar la energía de los fotones para transmitirla al pigmento diana; y el centro de reacción. Este está formado por proteínas y por pigmentos, encontrándose en él el llamado pigmento diana, que es aquel que recibe la energía de excitación de la antena, energía que sirve para excitar y liberar electrones. Aquí también se encuentra el primer dador de electrones, que repone los electrones al pigmento diana,
Primer aceptor, que recibe los electrones liberados.
Hay dos tipos de fotosistemas:
Fotosistema I, que se encuentra sobre todo en los tilacoides de estroma, y cuyo pigmento diana es la clorofila P700.
Fotosistema II, que se encuentra sobre todo en los grana y cuyo pigmento diana es la clorofila P680.
Proceso.
El proceso de la fase luminosa, supuesto para dos electrones, es el siguiente: Los fotones inciden sobre el fotosistema II, excitando y liberando dos electrones, que pasan al primer aceptor de electrones, la feofitina. Los electrones los repone el primer dador de electrones, el dador Z, con los electrones procedentes de la fotólisis del agua en el interior del tilacoide (la molécula de agua se divide en 2H+ + 2e- + 1/2O2). Los protones de la fotólisis se acumulan en el interior del tilacoide, y el oxígeno es liberado.
Los electrones pasan a una cadena de transporte de electrones, que invertirá su energía liberada en la síntesis de ATP. ¿Cómo? La teoría quimioosmótica nos lo explica de la siguiente manera: los electrones son cedidos a las plastoquinonas, las cuales captan también dos protones del estroma. Los electrones y los protones pasan al complejo de citocromos bf, que bombea los protones al interior del tilacoide. Se consigue así una gran concentración de protones en el tilacoide (entre éstos y los resultantes de la fotólisis del agua), que se compensa regresando al estroma a través de las proteínas ATP-sintasas, que invierten la energía del paso de los protones en sintetizar ATP. La síntesis de ATP en la fase fotoquímica se denomina fotofosforilación.
Los electrones de los citocromos pasan a la plastocianina, que los cede a su vez al fotosistema I. Con la energía de la luz, los electrones son de nuevo liberados y captados por el aceptor A0. De ahí pasan a través de una serie de filoquinonas hasta llegar a la ferredoxina. Ésta molécula los cede a la enzima NADP+-reductasa, que capta también dos protones del estroma. Con los dos protones y los dos electrones, reduce un NADP+ en NADPH + H+.
El balance final es: por cada molécula de agua (y por cada cuatro fotones) se forman media molécula de oxígeno, 1,3 moléculas de ATP, y un NADPH + H+.
Fase luminosa cíclica.
En la fase luminosa o fotoquímica cíclica interviene de forma exclusiva el fotosistema I, generándose un flujo o ciclo de electrones que en cada vuelta da lugar a síntesis de ATP. Al no intervenir el fotosistema II, no hay fotólisis del agua y, por ende, no se produce la reducción del NADP+ ni se desprende oxígeno. Únicamente se obtiene ATP.
El objetivo que tiene la fase cíclica tratada es el de subsanar el déficit de ATP obtenido en la fase acíclica para poder afrontar la fase oscura posterior.
Cuando se ilumina con luz de longitud de onda superior a 680 nm (lo que se llama rojo lejano) sólo se produce el proceso cíclico. Al incidir los fotones sobre el fotosistema I, la clorofila P700 libera los electrones que llegan a la ferredoxina, la cual los cede a un citocromo b6 y éste a la plastoquinona (PH), que capta dos protones y pasa a (PQH2). La plastoquinona reducida cede los dos electrones al citocromo f e introduce
Tiene lugar al mismo tiempo que la acíclica. En ella sólo interviene el fotosistema I. Los electrones liberados, después de llegar a la ferredoxina, pasan a las plastoquinonas, y siguen la cadena de transporte de electrones hasta regresar a la plastocianina y al fotosistema I. Por tanto, se genera ATP pero no NADPH. Sirve para compensar el hecho de que en la fotofosforilación acíclica no se genera suficiente ATP para la fase oscura.
BIBLIOGRAFÍA.
http://sisbib.unmsm.edu.pe/BVRevistas/dermatologia/v12_n2/fotobiologia_cutanea.htm
http://www.textoscientificos.com/fotografia/fotoquimica


